
On to the next of Tinbergen’s four questions!
As a quick reminder (since I am really dragging this series out now), I have previously defined consciousness very broadly. I posit that this still amorphous concept can best be understood as the set of processes where living organisms (governed by the various laws of selection) sense and respond to biological forces. This is currently only achieved by carbon-based life, but there’s no reason that artificial life couldn’t conceivably fulfill these criteria too.
I first described this definition in my brief history of everything that has ever existed. To fully grasp any biological phenomenon (which consciousness surely is in a natural view of the universe), Nikolaas Tinbergen developed a 2x2 matrix of things to consider, which has since become the standard in evolutionary studies. His four items are: 1) function (adaptation), 2) mechanism (causation), 3) ontogeny (development), and 4) phylogeny (evolution). In my last post, I covered the first of these items--the functions of consciousness—which led to the following hierarchical table:
As a quick reminder (since I am really dragging this series out now), I have previously defined consciousness very broadly. I posit that this still amorphous concept can best be understood as the set of processes where living organisms (governed by the various laws of selection) sense and respond to biological forces. This is currently only achieved by carbon-based life, but there’s no reason that artificial life couldn’t conceivably fulfill these criteria too.
I first described this definition in my brief history of everything that has ever existed. To fully grasp any biological phenomenon (which consciousness surely is in a natural view of the universe), Nikolaas Tinbergen developed a 2x2 matrix of things to consider, which has since become the standard in evolutionary studies. His four items are: 1) function (adaptation), 2) mechanism (causation), 3) ontogeny (development), and 4) phylogeny (evolution). In my last post, I covered the first of these items--the functions of consciousness—which led to the following hierarchical table:

Now that we are clear about all of the biological functions we are talking about in the multi-faceted and complex concept of consciousness, we can try to answer Tinbergen’s next three questions by looking for the current mechanisms and the personal and evolutionary histories that are associated with these functions. Let’s do the mechanisms first and see how they fit within the hierarchy shown above.
Just like last time, there are lot of intricate details for this large and unwieldy topic, so I’ll write simple numbered statements followed by their justifications so you can quickly read the statements to get the gist of the argument, or you can dip into any of the details you might want for further information (with links there to even more). Unlike last time, however, we now have a structure to work within, so I’ll try to abide by that. I should also note that it is impossible to cover the details of all of the mechanisms of all of the facets of consciousness for all of the living creatures that have ever experienced it. This post topped out at nearly 13,000 words when I included all of the details I had gathered to help me understand the mechanisms, but I have shortened it considerably (around 8,000 words now) to merely focus on the general gist of the types of mechanisms that are out there. I’m confident you can always find more from there on any specific mechanisms that interest you. Okay. Here we go!
1.0 Origin of Life (The first three criteria for life are: organisation, growth, and reproduction.)
1.01 Our current best guess for the origin of life involves lipid vesicles containing polymers that can grow and divide. The chemical structures describe the organisation. Osmotic pressure and new bonds caused growth. Mechanical forces split these growing vesicles, which led to reproduction and thus evolution.
1.02 These are the specific forms of early chemical life, but what defines them can be generalized into abstract terms. The border between any self and non-self has thus been defined as a Markov blanket. Markov blankets have three characteristics—a physical boundary, sensory systems on the boundary, and internal mechanisms that enable the self to exist and persist.
2.0 Affect (The first four cognitive abilities—response to stimuli, adaptation, homeostasis, and metabolism—enable the fulfillment of the final four criteria for life: sense perception, valence, discrimination, and motivation.)
2.01 For the earliest forms of biological life, changes to their molecular structures exert forces on those molecules. Being surrounded by stronger or weaker osmotic forces defines the presence of others. Identical osmotic forces indicate groups of the same entities. Growth is good for survival. Loss is bad for it. This is how the first subjects begin to sense change in their environments, assign valence to these changes, and discriminate between selves and not-selves.
2.02 Changes in the organizational structure of living molecules are due to the action potentials of ion flows across cell membranes. A few kinds of ions have evolved to be especially important in life’s current biochemistry for controlling basic stimulus-response mechanisms. These may be able to be mimicked in artificial life, but it would take much more diversity than is often considered.
2.03 Chemical building blocks provide the ability to process information, which enables the repeatable decisions (cognition) necessary to remain alive.
2.04 In abstract terminology, these particular chemical responses to physical stimuli are the processing of information within Markov Blankets. They are systematically selected for by the natural processes of evolution. Logically, the systems that survive are those that enable survival. The resulting stability is called homeostasis, and the chemical processes that sustain life are its metabolism.
2.05 Deviations from homeostasis cause internal reactions that are selected to bring systems back to their preferred state. These various reactions are the affective core of consciousness. These reactions diverged over the course of evolution into distinct facets that are recognizable as the seven basic emotions.
2.06 Note that the conscious awareness and processing of affective reactions comes later in the hierarchy of consciousness. Such cognition only rides on the affective core and cannot exist in biological systems on its own.
3.0 Intention (Five more cognitive abilities—attention, memory, pattern recognition, learning, and communication—enable intentional actions of the core self, eventually including the delay of reflexes.)
3.01 Core affect cannot be responded to by internal reactions alone—externally observable behavioral responses must occur as well. Once they do, it can be said that these organisms act with intention. They become driven.
3.02 Further evolution along this path produces further mechanisms for producing intentions. Once multicellular forms of life evolve sufficient complexity, hormones act for intercellular communication, which allows whole organisms to respond with intent towards naturally selected goals.
3.03 This evolution continues on to produce neurons in some forms of life. These are incredibly diverse and provide an enormous amount of additional abilities, including internally generated actions.
3.04 Neurons proliferate and form nervous systems in animals that enable further reactions to the environment.
3.05 Nervous systems come together into nodes that evolve into more and more sophisticated brains. These are used to coordinate multiple streams of sensory information.
3.06 Evolutionary pressures on organisms led them to develop brain modules, systems, and networks, which work in nested hierarchies to enable more and more complexity and effectiveness in understanding and responding to the world.
Just like last time, there are lot of intricate details for this large and unwieldy topic, so I’ll write simple numbered statements followed by their justifications so you can quickly read the statements to get the gist of the argument, or you can dip into any of the details you might want for further information (with links there to even more). Unlike last time, however, we now have a structure to work within, so I’ll try to abide by that. I should also note that it is impossible to cover the details of all of the mechanisms of all of the facets of consciousness for all of the living creatures that have ever experienced it. This post topped out at nearly 13,000 words when I included all of the details I had gathered to help me understand the mechanisms, but I have shortened it considerably (around 8,000 words now) to merely focus on the general gist of the types of mechanisms that are out there. I’m confident you can always find more from there on any specific mechanisms that interest you. Okay. Here we go!
1.0 Origin of Life (The first three criteria for life are: organisation, growth, and reproduction.)
1.01 Our current best guess for the origin of life involves lipid vesicles containing polymers that can grow and divide. The chemical structures describe the organisation. Osmotic pressure and new bonds caused growth. Mechanical forces split these growing vesicles, which led to reproduction and thus evolution.
- Let’s review: Monomers diffuse into a fatty acid vesicle. Monomers spontaneously polymerize and copy any template. Heat separates strands and increases membrane permeability to monomers. Polymer backbones attract ions, increasing osmotic pressure. Pressure on the membrane drives its growth at the expense of nearby vesicles containing less polymer. Vesicles grow into tubular structures. Mechanical forces cause vesicles to divide. Daughter vesicles inherit polymers from the parent vesicle. Polymer sequences that replicate faster will dominate the population. Thus beginning evolution! (Post 17)
1.02 These are the specific forms of early chemical life, but what defines them can be generalized into abstract terms. The border between any self and non-self has thus been defined as a Markov blanket. Markov blankets have three characteristics—a physical boundary, sensory systems on the boundary, and internal mechanisms that enable the self to exist and persist.
- For a system to resist entropy, three conditions must be met: (i) There must be a boundary which separates the internal and external states of the system, and thereby insulates the system from the world. Let's call the former states “the system” and the latter states “the not-system.” (ii) There must be a mechanism which registers the influence of dissipative external forces—i.e. the free energy. Let's call this mechanism the “sensory states” of the system. (iii) There must be a mechanism which counteracts these dissipative forces—i.e. which binds the free energy. Let's call this mechanism the “active states” of the system, such as motor and autonomic reflexes. … According to Friston (2013), these functional conditions—which enable self-organizing systems to exist and persist over time—emerge naturally (indeed necessarily) within any ergodic random dynamical system that possesses a Markov blanket. This blanket establishes the boundary conditions above. (Solms)
2.0 Affect (The first four cognitive abilities—response to stimuli, adaptation, homeostasis, and metabolism—enable the fulfillment of the final four criteria for life: sense perception, valence, discrimination, and motivation.)
2.01 For the earliest forms of biological life, changes to their molecular structures exert forces on those molecules. Being surrounded by stronger or weaker osmotic forces defines the presence of others. Identical osmotic forces indicate groups of the same entities. Growth is good for survival. Loss is bad for it. This is how the first subjects begin to sense change in their environments, assign valence to these changes, and discriminate between selves and not-selves.
- Molecules are held together by covalent bonds, which involve the sharing of electron pairs between atoms. … Intermolecular forces are the forces which mediate interactions between molecules and other types of neighboring particles such as atoms or ions. … The four key intermolecular forces are: 1) Ionic bonds; 2) Hydrogen bonding; 3) Van der Waals dipole-dipole interactions; and 4) Van der Waals dispersion forces. (Post 17)
- Any changes to biological molecules would generate forces. These forces are exerted on singularly identifiable objects. (Post 19)
- I propose that these chemical forces, once in service of biological needs, are the defined starting point for turning objects into subjects. (Post 19)
2.02 Changes in the organizational structure of living molecules are due to the action potentials of ion flows across cell membranes. A few kinds of ions have evolved to be especially important in life’s current biochemistry for controlling basic stimulus-response mechanisms. These may be able to be mimicked in artificial life, but it would take much more diversity than is often considered.
- Membrane potential is the difference in electric potential between the interior and the exterior of a biological cell. All animal cells are surrounded by a membrane composed of a lipid bilayer with proteins embedded in it. The membrane serves as both an insulator and a diffusion barrier to the movement of ions. Transmembrane proteins, also known as ion transporter or ion pump proteins, actively push ions across the membrane and establish concentration gradients across the membrane, and ion channels allow ions to move across the membrane down those concentration gradients. Ion pumps and ion channels are electrically equivalent to a set of batteries and resistors inserted in the membrane, and therefore create a voltage between the two sides of the membrane. The membrane potential has two basic functions. First, it allows a cell to function as a battery, providing power to operate a variety of “molecular devices” embedded in the membrane. Second, in electrically excitable cells such as neurons and muscle cells, it is used for transmitting signals between different parts of a cell. (Membrane potential)
- In physiology, an action potential occurs when the membrane potential of a specific cell location rapidly rises and falls: this depolarization then causes adjacent locations to similarly depolarize. Action potentials occur in some plant cells and in several types of animal cells, called excitable cells, which include neurons, muscle cells, endocrine cells, and glomus cells. (Action potential)
- Voltage-gated-calcium-channels (VGCCs) are normally closed. They are activated (i.e., opened) at depolarized membrane potentials. The concentration of calcium (Ca2+ ions) is normally several thousand times higher outside the cell than inside. Activation of particular VGCCs allows a Ca2+ influx into the cell, which, depending on the cell type, results in activation of calcium-sensitive potassium channels, muscular contraction, excitation of neurons, up-regulation of gene expression, or release of hormones or neurotransmitters. (Voltage-gated Calcium Channel)
- Voltage-gated sodium channels play an important role in action potentials. If enough channels open when there is a change in the cell's membrane potential, a small but significant number of Na+ ions will move into the cell down their electrochemical gradient, further depolarizing the cell. Thus, the more Na+ channels localized in a region of a cell's membrane the faster the action potential will propagate and the more excitable that area of the cell will be. The ability of these channels to assume a closed-inactivated state causes the refractory period and is critical for the propagation of action potentials down an axon. (Sodium Channel)
- A refractory period is a period of time during which an organ or cell is incapable of repeating a particular action, or (more precisely) the amount of time it takes for an excitable membrane to be ready for a second stimulus once it returns to its resting state following an excitation. It most commonly refers to electrically excitable muscle cells or neurons. (Refractory period)
- There are about 100 billion neurons in the human brain, each of which forms synapses with many other neurons. A synapse is the gap between two neurons (known as the presynaptic and postsynaptic neurons). The presynaptic neuron releases neurotransmitters, such as glutamate and GABA, which bind to receptors on the postsynaptic cell membrane, activating ion channels. Opening and closing those channels changes the cell's electrical potential. If the potential changes dramatically enough, the cell fires an electrical impulse called an action potential. (Mimicking the Brain in Silicon)
- MIT researchers designed a computer chip so that the transistors could mimic the activity of different ion channels. While most chips operate in a binary, on/off mode, current flows through the transistors on the new brain chip in analogue, not digital, fashion. A gradient of electrical potential drives current to flow through the transistors just as ions flow through ion channels in a cell. (Mimicking the Brain in Silicon)
- This synapse diversity could have implications for the prospect of creating artificial consciousness. (Not intelligence, but consciousness.) No computer has synapse diversity. I have met numerous people who are experts in building computers based on neural principles. As interesting as I find their presentations, they typically have built them on principles that are several decades out of date. There’s no concept of the molecular organization. It’s based on a few electrophysiological parameters that are known about neurons. which comes from the era of cells and electrophysiology. Those are sort of pre-1990’s stuff. (Seth Grant)
2.03 Chemical building blocks provide the ability to process information, which enables the repeatable decisions (cognition) necessary to remain alive.
- A candidate mechanism that may serve as the biological basis of the continuum of cognitive function [is] the chemistry of protein networks, whose potential information-processing power and similarity to neural networks in single cells was first described by Cambridge zoologist Dennis Bray, who noticed that “many proteins in living cells appear to have as their primary function the transfer and processing of information, rather than the chemical transformation of metabolic intermediates or the building of cellular structure.” (Lyon)
- Protein signal transduction networks should be considered the basis of cognitive function. Neurons and the electrical properties of neurons come first to mind in discussions of the brain, but chemical protein networks are also widely used in that organ because, energetically, they are the cheapest way of sending and receiving information. (Lyon)
- In biological systems, information is transmitted “whenever a source’s change in state registers as a change in state at a receiver.” The change may be in environmental pH; the availability of nutrients; chemical indicators of conspecifics, predators, or prey; build-up of reactive oxygen species; osmolarity; diffusion potential, and so on. (Lyon)
2.04 In abstract terminology, these particular chemical responses to physical stimuli are the processing of information within Markov Blankets. They are systematically selected for by the natural processes of evolution. Logically, the systems that survive are those that enable survival. The resulting stability is called homeostasis, and the chemical processes that sustain life are its metabolism.
- For self-organizing systems—including all living things, like us—to exist, they must resist entropy. That is, self-organizing systems can only persist over time by occupying “preferred” states—as opposed to being dispersed over all possible states, and thereby dissipating. (Solms)
- A Markov blanket can only “know” states of the not-system vicariously. In other words, external states can only be “inferred” by the system on the basis of “sensory impressions” upon the Markov blanket. (Solms)
- In summary, homeostasis is explained by the causal dynamics mandated by the very existence of Markov blankets; in terms of which self-organizing systems generate a type of work that binds free energy and maintains the system in its typically occupied (“preferred” or “valued”) states. (Solms)
- Metabolism is the set of life-sustaining chemical reactions in organisms. The three main purposes of metabolism are: the conversion of food to energy to run cellular processes; the conversion of food/fuel to building blocks for proteins, lipids, nucleic acids, and some carbohydrates; and the elimination of metabolic wastes. These enzyme-catalyzed reactions allow organisms to grow and reproduce, maintain their structures, and respond to their environments. (Metabolism)
2.05 Deviations from homeostasis cause internal reactions that are selected to bring systems back to their preferred state. These various reactions are the affective core of consciousness. These reactions diverged over the course of evolution into distinct facets that are recognizable as the seven basic emotions.
- [There] are various instinctual motivational circuits. These are also known as the circuits for “basic emotion”. There are several classifications of these emotions. The best-known examples are those that generate (1) appetitive foraging, (2) consummatory reward, (3) freezing and flight, (4) aggressive attack, (5) nurturant care, (6) separation distress, and (7) rough-and-tumble play. It is important to note that each of the instinctual circuits generates not only stereotyped behaviors but also diverse feeling states, such as expectant interest, orgasmic delight, trepidatious fear, destructive rage, loving affection, sorrowful grief, and exuberant joy. The circuits for these basic emotions are conserved across the mammalian series, and they admit of considerable chemical specificity. They are no less innate than the vital evolutionary survival and sexual needs which gave rise to them. They are unconditioned “tools for living”. (Solms and Panksepp)
2.06 Note that the conscious awareness and processing of affective reactions comes later in the hierarchy of consciousness. Such cognition only rides on the affective core and cannot exist in biological systems on its own.
- The removal of the neocortex has long been known to spare emotionality. Indeed, not only are the rewarding effects of subcortical brain stimulations demonstrably preserved in decorticated creatures, these animals are actually more emotional than normal. The most strikingly concordant human evidence to emerge in recent years, relevant to this broader question, concerns a condition called hydranencephaly, in which the cerebral cortex as a whole is destroyed in utero. However, the subcortical networks are functional; thus, the children are markedly emotionally functional human beings. “They express pleasure by smiling and laughter, and aversion by ‘fussing’ arching of the back and crying (in many gradations), their faces being animated by these emotional states. A familiar adult can employ this responsiveness to build up play sequences predictably progressing from smiling, through giggling, to laughter and great excitement on the part of the child.” They also show basic emotional learning. Although there is in these children significant degradation of the types of consciousness that are normally associated with external perception, there can be no doubt that they are conscious, both quantitatively and qualitatively. (Solms and Panksepp)
- Let us consider a third problem with the cortico-centric approach. The third problem is that there is a brain structure which does pass the critical test just mentioned. This structure is located not in the cortex but the brainstem. Consciousness is obliterated by focal lesions of the brainstem core—in a region conventionally described as the extended reticulothalamic activating system (ERTAS). Recent findings indicate that the smallest lesions within the brainstem which cause total loss of consciousness (i.e., coma) are located in or near the parabrachial nuclei of the pons. (Solms)
- Although many cognitive scientists still must be weaned of the view that the cerebral cortex is the seat of consciousness, the weight of evidence for the alternative view that the arousal processes generated in the upper brainstem and limbic system feel like something in and of themselves, is now overwhelming. (Solms)
- This conclusion is further supported by the fact that drugs acting on the neuromodulators sourced in the ERTAS nuclei (serotonin, dopamine, noradrenaline, acetylcholine) have powerful effects on mood and anxiety, etc.—which is why they represent the mainstay of psychopharmacology today. (Solms)
3.0 Intention (Five more cognitive abilities—attention, memory, pattern recognition, learning, and communication—enable intentional actions of the core self, eventually including the delay of reflexes.)
3.01 Core affect cannot be responded to by internal reactions alone—externally observable behavioral responses must occur as well. Once they do, it can be said that these organisms act with intention. They become driven.
- The dynamics of a Markov blanket generate two fundamental properties—namely (elemental forms of) selfhood and intentionality. [T]hese dynamics also generate elemental properties of bodies—namely an insulating membrane and adaptive behavior. (Solms)
- Body-monitoring nuclei…can only go so far in terms of meeting endogenous needs through internal (autonomic) adjustments. Beyond that limit, external action is called for. At that point, autonomic reflexes become drives. [For example], interoceptive “need detectors” trigger not only autonomic reflexes but also feelings of hunger, thirst, etc. These drives typically trigger “foraging” behaviors (“SEEKING”). (Solms)
3.02 Further evolution along this path produces further mechanisms for producing intentions. Once multicellular forms of life evolve sufficient complexity, hormones act for intercellular communication, which allows whole organisms to respond with intent towards naturally selected goals.
- At the most basic level, the function of the nervous system is to send signals from one cell to others, or from one part of the body to others. There are multiple ways that a cell can send signals to other cells. One is by releasing chemicals called hormones into the internal circulation, so that they can diffuse to distant sites. (Nervous Systems)
- A hormone is any member of a class of signaling molecules, produced by glands in multicellular organisms, that are transported by the circulatory system to target distant organs to regulate physiology and behavior. The glands that secrete hormones comprise the endocrine signaling system. Hormones affect distant cells by binding to specific receptor proteins in the target cell, resulting in a change in cell function. (Hormones)
- Hormones serve to communicate between organs and tissues for physiological regulation and behavioral activities such as digestion, metabolism, respiration, tissue function, sensory perception, sleep, excretion, lactation, stress induction, growth and development, movement, reproduction, and mood manipulation. (Hormones)
3.03 This evolution continues on to produce neurons in some forms of life. These are incredibly diverse and provide an enormous amount of additional abilities, including internally generated actions.
- In contrast to the “broadcast” mode of hormone signaling, the nervous system provides “point-to-point” signals—neurons project their axons to specific target areas and make synaptic connections with specific target cells. Thus, neural signaling is capable of a much higher level of specificity than hormonal signaling. It is also much faster: the fastest nerve signals travel at speeds that exceed 100 meters per second. (Nervous Systems)
- A cell that receives a synaptic signal from a neuron may be excited, inhibited, or otherwise modulated. (Nervous Systems)
- Even in the nervous system of a single species such as humans, hundreds of different types of neurons exist, with a wide variety of morphologies and functions. These include sensory neurons that transmute physical stimuli such as light and sound into neural signals, and motor neurons that transmute neural signals into activation of muscles or glands. (Nervous Systems)
- There are literally hundreds of different types of synapses. In fact, there are over a hundred known neurotransmitters, and many of them have multiple types of receptors. Molecular neuroscientists generally divide receptors into two broad groups: chemically gated ion channels and second messenger systems. When a second messenger system is activated, it starts a cascade of molecular interactions inside the target cell, which may ultimately produce a wide variety of complex effects, such as increasing or decreasing the sensitivity of the cell to stimuli, or even altering gene transcription. (Nervous Systems)
- Over the 1990’s, there was a general model of synapses which was that they contained very few proteins and those few proteins that were known, which you could count on one hand more or less, were sufficient to produce synaptic transmission and synaptic plasticity. People thought that could account for learning, but it wasn’t like that at all. What we found was that on the post-synaptic side of synapses (the side of the synapse where information first comes into a nerve cell), we identified ten times the number of proteins that were previously known, which suddenly revealed a very much unexpected complexity. Quite a lot of people at the time thought that it was some sort of artifact, but it wasn’t. It was actually only 1/10th as complex as what it turned out to be! Over the subsequent years, we found another tenfold more proteins. Many labs have confirmed this now. Essentially, inside a synapse, on the post-synaptic side, you can have more than a thousand types of proteins in the synapse. This really changed the way of thinking, from the synapse being just a “connector” in the nervous system. That’s not a very nice way to talk about a synapse because in fact it’s a super-sophisticated molecular computer. (Seth Grant)
- The human brain has a vast number of synapses—somewhere on the order of a million billion of them. So, we have a vast number of synapses, and we have a large number of proteins in the synapses. We have also uncovered evidence that these synapse proteins are not distributed the same across all synapses. In some parts of the brain, some proteins are found. In other parts of the brain, other proteins are found. And that was giving us this clue that there was this synapse diversity at a level that we hadn’t really thought of before. (Seth Grant)
- In 2018, we published a paper that had been the first brain-wide survey of synapse diversity using a variety of methods. We now call this diversity the “synaptome.” In the way the “genome” is all of the genes that an animal has, the synaptome is all of the synapses that an animal has. Just as genes have an architecture, synapse diversity has an architecture. We find that different types of synapses are found in different parts of the brain and they have certain proteins. Interestingly, those parts of the brain that are involved in higher cognitive functions—in the cortex and the hippocampus—are the parts where you find the most diversity of the types of synapses. This is telling us something important. Having all of those types of synapses is probably giving very sophisticated computation to the brain circuits for the types of behavior like language, speech, and memory processing. (Seth Grant)
- If you look at the potential of the different combinations of these proteins, it’s very easy to imagine that every single synapse could be different. A mouse has about 10 to the power of 11 synapses. Even with a small number of proteins, say 10, it’s possible to have every one of those synapses be different. But as I’ve already said, there are more than a thousand in there and they also have other post-translation modifications. So, it would be very simple to have a mouse brain where every synapse is actually different. It could also easily be the case that in the human brain, which is much bigger, every synapse could be different. We don’t think that’s going to be the case. We think they are going to be organized and there are going to be abundant classes and non-abundant classes, and some might even have redundant functions meaning they might have different molecular makeups but function the same way. But there is plenty of scope for what you might call species differences in the synapse composition. I wouldn’t be at all surprised if there were some types of synapses that are unique to mice, and some that are unique to humans. There will also be some that are conserved between the two species. It’s going to be crucial to look at that. (Seth Grant)
- A neuron is called “identified” if it has properties that distinguish it from every other neuron in the same animal and if every individual organism belonging to the same species has one and only one neuron with the same set of properties. In vertebrate nervous systems, very few neurons are “identified” in this sense—in humans, there are believed to be none—but in simpler nervous systems, some or all neurons may be thus unique. In the roundworm C. elegans, whose nervous system is the most thoroughly described of any animal's, every neuron in the body is uniquely identifiable. One notable consequence of this fact is that the form of the C. elegans nervous system is completely specified by the genome, with no experience-dependent plasticity. (Nervous Systems)
- One very important subset of synapses is capable of forming memory traces by means of long-lasting activity-dependent changes in synaptic strength. The best-known form of neural memory is a process called long-term potentiation (LTP), which operates at synapses that use the neurotransmitter glutamate acting on a special type of receptor known as the NMDA receptor. Since the discovery of LTP in 1973, many other types of synaptic memory traces have been found, involving increases or decreases in synaptic strength that are induced by varying conditions, and last for variable periods of time. (Nervous Systems)
- Because of the variety of voltage-sensitive ion channels that can be embedded in the membrane of a neuron, many types of neurons are capable, even in isolation, of generating rhythmic sequences of action potentials, or rhythmic alternations between high-rate bursting and quiescence. When neurons that are intrinsically rhythmic are connected to each other by excitatory or inhibitory synapses, the resulting networks are capable of a wide variety of dynamical behaviors. (Nervous Systems)
3.04 Neurons proliferate and form nervous systems in animals that enable further reactions to the environment.
- The connections between neurons can form neural pathways, neural circuits, and larger networks that generate an organism's perception of the world and determine its behavior. (Nervous Systems)
- The basic neuronal function of sending signals to other cells includes a capability for neurons to exchange signals with each other. Networks formed by interconnected groups of neurons are capable of a wide variety of functions, including feature detection, pattern generation, and timing, and there are seen to be countless types of information processing possible. (Nervous Systems)
- The nervous system is a highly complex part of an animal that coordinates its actions and sensory information by transmitting signals to and from different parts of its body. In vertebrates it consists of two main parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain and spinal cord. The PNS consists mainly of nerves, which are enclosed bundles of the long fibers or axons, that connect the CNS to every other part of the body. The PNS is divided into three separate subsystems, the somatic, autonomic, and enteric nervous systems. Somatic nerves mediate voluntary movement. The autonomic nervous system is further subdivided into the sympathetic and the parasympathetic nervous systems. The sympathetic nervous system is activated in cases of emergencies to mobilize energy, while the parasympathetic nervous system is activated when organisms are in a relaxed state. The enteric nervous system functions to control the gastrointestinal system. Both autonomic and enteric nervous systems function involuntarily. (Nervous Systems)
3.05 Nervous systems come together into nodes that evolve into more and more sophisticated brains. These are used to coordinate multiple streams of sensory information.
- In many species, the great majority of neurons participate in the formation of centralized structures (the brain and ganglia) and they receive all of their input from other neurons and send their output to other neurons. (Nervous Systems)
- Nervous systems are found in most multicellular animals but vary greatly in complexity. The only multicellular animals that have no nervous system at all are sponges, placozoans, and mesozoans, which have very simple body plans. The nervous systems of the radially symmetric organisms ctenophores (comb jellies) and cnidarians (which include anemones, hydras, corals and jellyfish) consist of a diffuse nerve net. All other animal species, with the exception of a few types of worm, have a nervous system containing a brain, a central cord (or two cords running in parallel), and nerves radiating from the brain and central cord. The size of the nervous system ranges from a few hundred cells in the simplest worms, to around 300 billion cells in African elephants. (Nervous Systems)
3.06 Evolutionary pressures on organisms led them to develop brain modules, systems, and networks, which work in nested hierarchies to enable more and more complexity and effectiveness in understanding and responding to the world.
- The cerebral cortex is the largest site of neural integration in the central nervous system. It plays a key role in attention, perception, awareness, thought, memory, language, and consciousness. (Cerebral Cortex)
- The Global Neuronal Workspace Theory (from Dehaene) is an intellectual descendent of the Global Workspace Theory (Baars). These theories identify brain modules for: balance and coordination; memory; emotion; language; writing; attention, planning, organization, reasoning; emotional affect, adaptability; motor / sensory; listening and decoding; reading and interpretation; visual-spatial, visual recognition. (Jerry Fodor called these modules informationally encapsulated, meaning somewhat private within each module.) … For some functions, there may be specific pathways through these modules, e.g. dorsal visual stream. … For general connections between multiple modules there may be a global workspace. This coordinates inputs from evaluative systems (value), attentional systems (focusing), long-term memory (past), and perceptual systems (present), into motor control outputs (future). Information in the global workspace is available from all modules and can be seen by each module. (Introduction to Brain Consciousness)
- The default mode network (DMN) is active when we’re internally focused, thinking about ourselves and using our memory and imagination. The dorsal attention network (DAT), on the other hand, is activated when we’re aware of and paying attention to the environment around us. (Ramirez)
- A team from the University of Michigan described their finding that the default mode network (DMN) and the dorsal attention network (DAT) are anti-correlated, meaning that when one is active, the other is suppressed. The team also found that neither network was highly active in people who were unconscious. These findings suggest that the interplay of the DMN and the DAT support consciousness by allowing us to interact with our surroundings then to quickly internalize those interactions, essentially turning our experiences into thoughts and memories. (Ramirez)
- Figure 1: (Solms and Panksepp)
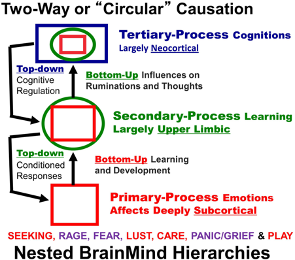
3.07 In summary, all of these chemical, neuronal, and brain developments have evolved to produce a variety of mechanisms (too numerous to list here) for driving intentional behaviour. This extends from the simplest forms of cognition in plants up to and including the most sophisticated varieties that we are aware of in humans. These mechanisms produce the expanding abilities of functions in the 3rd level of my hierarchy of consciousness (attention, memory, pattern recognition, learning, and communication).
- Plant cognition is a field of research directed at experimentally testing the cognitive abilities of plants, including perception, learning processes, memory, and consciousness. Although they lack a brain and the function of a conscious working nervous system, plants are still somehow capable of being able to adapt to their environment and change the integration pathway that would ultimately lead to how a plant “decides” to take response to a presented stimulus. (Plant Cognition)
- A plant known as the Mimosa pudica was tested for the ability to adapt to closing its leaves upon repeated drops with no apparent harm appointed to the plant. The results showed that with repeated drops, the Mimosa pudica eventually stopped closing its leaves or opened its leaves quicker. This behavior exhibited a trait in which the plant has adapted to not closing, or showing minimal closing, when repeated exposure to a non-harming situation is coupled with its own defence behavior. (Plant Cognition)
- We declare the following: “The absence of a neocortex does not appear to preclude an organism from experiencing affective states. Convergent evidence indicates that non-human animals have the neuroanatomical, neurochemical, and neurophysiological substrates of conscious states along with the capacity to exhibit intentional behaviors. Consequently, the weight of evidence indicates that humans are not unique in possessing the neurological substrates that generate consciousness. Non-human animals, including all mammals and birds, and many other creatures, including octopuses, also possess these neurological substrates.” (Cambridge Declaration of Consciousness)
- Neural circuits supporting behavioral / electrophysiological states of attentiveness, sleep, and decision making appear to have arisen in evolution as early as the invertebrate radiation, being evident in insects and cephalopod molluscs (e.g., octopus). (Cambridge Declaration of Consciousness)
- Present-tense emotionality is what communication by animals is mostly about. (Sapolsky)
- Joint attention refers to when two people look at and attend to the same thing; parents often use the act of pointing to prompt infants to engage in joint attention. The inclination to spontaneously reference an object in the world as of interest, via pointing, and to likewise appreciate the directed attention of another, may be the underlying motive behind all human communication. (Theory of Mind Wikipedia)
3.08 Note that all of these responses to stimuli are initially observable as reflexes. Later, brain modules and networks evolved that could sense and respond to other parts of the brain, as well as the internal affective moods that influence the entire organism. Such “meta” modules and networks are able to gain control over simpler systems by interrupting their pathways to action. This allows delays to what would otherwise be viewed as automatic responses. In effect, these act as internal (and therefore invisible) reflexes where logical if-then statements control local behavior based on the wider context that is reported from other senses or memories.
- The simplest type of neural circuit is a reflex arc, which begins with a sensory input and ends with a motor output, passing through a sequence of neurons connected in series. This can be shown in the “withdrawal reflex” causing a hand to jerk back after a hot stove is touched. (Nervous Systems)
- Although the simplest reflexes may be mediated by circuits lying entirely within the spinal cord, more complex responses rely on signal processing in the brain. For example, when an object in the periphery of the visual field moves, and a person looks toward it, many stages of signal processing are initiated. The initial sensory response, in the retina of the eye, and the final motor response, in the oculomotor nuclei of the brain stem, are not all that different from those in a simple reflex, but the intermediate stages are completely different. Instead of a one- or two-step chain of processing, the visual signals pass through perhaps a dozen stages of integration, involving the thalamus, cerebral cortex, basal ganglia, superior colliculus, cerebellum, and several brainstem nuclei. These areas perform signal-processing functions that include feature detection, perceptual analysis, memory recall, decision-making, and motor planning. (Nervous Systems)
4.0 Prediction (This level in the hierarchy of consciousness is enabled by mechanisms for the cognitive abilities of anticipation, problem solving, and error detection.)
4.01 Once the mechanisms for memory, pattern recognition, and learning are set in place by the 3rd level of consciousness, animals with brains can begin to analyze current situations in order to predict what will happen next. This ability provides an enormous advantage to those who can master it.
- We can only perceive one signal at a time. And there is a 1/3 second time lag. Error prediction makes up for this. (Post 9)
4.02 There are various internal mechanisms that enable prediction. These fall under the general banner of predictive coding and include anticipation, problem solving, error detection, and feelings of precision.
- Predictive coding is a theory of brain function in which the brain is constantly generating and updating a mental model of the environment. The model is used to generate predictions of sensory input that are compared to actual sensory input. This comparison results in prediction errors that are then used to update and revise the mental model. (Predictive Coding)
- The understanding of perception as the interaction between sensory stimuli (bottom-up) and conceptual knowledge (top-down) continued to be established by Jerome Bruner who, starting in the 1940s, studied the ways in which needs, motivations, and expectations influence perception, which came to be known as 'New Look' psychology. (Predictive Coding)
- The brain solves the seemingly intractable problem of modeling distal causes of sensory input through a version of Bayesian inference. It does this by modeling predictions of lower-level sensory inputs via backward connections from relatively higher levels in a cortical hierarchy. Constrained by the statistical regularities of the outside world (and certain evolutionarily prepared predictions), the brain encodes top-down generative models at various temporal and spatial scales in order to predict and effectively suppress sensory inputs rising up from lower levels. A comparison between predictions and sensory input yields a difference measure which, if it is sufficiently large beyond the levels of expected statistical noise, will cause the generative model to update so that it better predicts sensory input in the future. (Predictive Coding)
- The neural evidence for this is still in its infancy. The empirical evidence for predictive coding is most robust for perceptual processing. (Predictive Coding)
- The anterior cingulate is heavily involved in “error detection,” noting discrepancies between what is anticipated and what occurs. (Sapolsky)
- Physiologically, precision is usually associated with the postsynaptic gain of cortical neurons reporting prediction errors. This is precisely the function of ERTAS modulatory neurons. (Solms)
4.03 Predictive coding changes living beings. They are no longer simply responders in the present tense to internal drives and external stimuli. Predictive beings are internally active thinkers trying to peer further and further into the future.
- Predictive coding inverts the conventional view of perception as a mostly bottom-up process, suggesting that it is largely constrained by prior predictions, where signals from the external world only shape perception to the extent that they are propagated up the cortical hierarchy in the form of prediction error. (Predictive Coding)
- Given that the world we live in is loaded with statistical noise, precision expectations must be represented as part of the brain's generative models, and they should be able to flexibly adapt to changing contexts. For instance, the expected precision of visual prediction errors likely varies between dawn and dusk, such that greater conditional confidence is assigned to errors in broad daylight than errors in prediction at nightfall. (Predictive Coding)
5.0 Awareness (This level in the hierarchy of consciousness is enabled by mechanisms for the cognitive abilities of self-reference.)
5.01 Through a mixture of abilities some brain networks gain general awareness of some parts of the self.
- [At first,] the external body is not a subject but an object, and it is perceived in the same register as other objects. Something has to be added to simple perception before one’s own body is differentiated from others. This level of representation (a.k.a. higher-order thought) enables the subject of consciousness to separate itself as an object from other objects. We envisage the process involving three levels of experience: (a) the subjective or phenomenal level of the anoetic self as affect, a.k.a. first-person perspective; (b) the perceptual or representational level of the noetic self as an object, no different from other objects, a.k.a. second-person perspective; (c) the conceptual or re-representational level of the autonoetic self in relation to other objects, i.e., perceived from an external perspective, a.k.a. third-person perspective. The self of everyday cognition is therefore largely an abstraction. That is why the self is so effortlessly able to think about itself in relation to objects, in such everyday situations as “I am currently experiencing myself looking at an object.” (Solms and Panksepp)
5.02 The mechanisms underlying this awareness may be specific to each species. This is a keen area of research, but they appear to just be more and more neuroanatomy.
- Another approach to studying consciousness applies specifically to the study of self-awareness, that is, the ability to distinguish oneself from others. The classic example of testing this is known as the mirror test, which involves placing a spot of coloring on the skin or fur near an individual's forehead and seeing if they attempt to remove it or at least touch the spot. Humans (older than 18 months) and other great apes, bottlenose dolphins, killer whales, pigeons, European magpies, and elephants have all been observed to pass this test. (Consciousness)
- Birds appear to offer, in their behavior, neurophysiology, and neuroanatomy a striking case of parallel evolution of consciousness. Certain species of birds have been found to exhibit neural sleep patterns similar to those of mammals, including REM sleep and, as was demonstrated in zebra finches, neurophysiological patterns, previously thought to require a mammalian neocortex. Magpies in particular have been shown to exhibit striking similarities to humans, great apes, dolphins, and elephants in studies of mirror self-recognition. Evidence of near human-like levels of consciousness has been most dramatically observed in African grey parrots. (Cambridge Declaration of Consciousness)
- When stimuli are presented to patients, but masked so they can’t detect it consciously, the visual cortex and amygdala are activated and that’s it. When the stimulus is not masked, you get activation in the visual cortex, the amygdala, and the prefrontal cortex as well. ... In order to be conscious of an apple, it not only needs to be represented in your visual cortex, it needs to be re-represented, which involves the prefrontal cortex. … So, the prefrontal cortex is emerging as an important area in the consolidation of our conscious experiences into what they are. (Post 12)
- When conscious access occurs, brain activity is strongly activated when a threshold of awareness is crossed. At that point the signal spreads to many brain areas. There are four highly reproducible signals associated with this. Signature 1: activation in parietal and prefrontal circuits. Signature 2: a slow wave called P3 that pairs late, approximately 1/3 second after stimulus (i.e. consciousness lags behind the world). Signature 3: deep brain electrodes detect late and sudden bursts of high frequency oscillations. Signature 4: information exchange across distant brain areas. (Post 9)
6.0 Abstraction (This level in the hierarchy of consciousness is enabled by mechanisms for understanding and creating symbols, art, language, memes, writing, mathematics, philosophy, and science, which all act to expand culture.)
6.01 Once the abstraction of self-awareness is established, further abstractions can also be made. Abstract thinking requires representing something in the brain that does not exist in front of it. Therefore, studying the mechanisms that underlie these abstract representations requires the self-report that is currently only available in humans. This is another area of ongoing research, but it appears that many brain areas are associated with distinct forms of abstract thinking.
- The gold standard for whether a response is conscious or not is whether you can talk about it. This doesn’t mean language and consciousness are identical, just that you have access to the experience to think about it (and we use language to discuss that access with one another). In non-human animal research, that doesn’t exist. (Post 12)
- Brodmann areas have been discussed, debated, refined, and renamed exhaustively for nearly a century and remain the most widely known and frequently cited cytoarchitectural organization of the human cortex. (Brodmann area)
- Many of the brain areas defined by Brodmann have their own complex internal structures. In a number of cases, brain areas are organized into topographic maps, where adjoining bits of the cortex correspond to adjoining parts of the body, or of some more abstract entity. (Brodmann area)
- The Broca area is a region in the frontal lobe of the dominant hemisphere of the brain (usually the left) with functions linked to speech production. (Broca’s area)
- Wernicke's area is one of the two parts of the cerebral cortex that are linked to speech, the other being Broca's area. It is involved in the comprehension of written and spoken language, in contrast to Broca's area, which is involved in the production of language. It is traditionally thought to reside in Brodmann area 22. (Wernicke’s area)
- Brodmann area 47 has been implicated in the processing of syntax in oral and sign languages, musical syntax, and semantic aspects of language. (Brodmann area)
- Higher order functions of the associated cortical areas are consistently localized to the same Brodmann areas by neurophysiological, functional imaging, and other methods. However, functional imaging can only identify the approximate localization of brain activations in terms of Brodmann areas since their actual boundaries in any individual brain requires its histological [post-mortem] examination. (Brodmann area)
6.02 These abstract abilities are useful for grasping all sorts of knowledge about the world. But they also gave humans greater moral capacities for responding to the world. For example, it would be very difficult for evolution to suddenly produce new emotional affects for the feelings of empathy and moral disgust, but human brains have learned to apply old reactions to new circumstances according to abstract rules. This greatly extended the flexibility of human consciousness.
- There are still ways that humans appear to stand alone. One of those is hugely important: the human capacity to think symbolically. Metaphors, similes, parables, figures of speech—they exert enormous power over us. (Sapolsky)
- As our hominid ancestors kept getting better at [abstract thinking], great individual and social advantages accrued. We became capable of representing emotions in the past and possible emotions in the future, as well as things that have nothing to do with emotion. We evolved a uniquely dramatic means of separating message from meaning and intent: lying. And we invented aesthetic symbolism; after all, those 30,000-year-old paintings of horses in Chauvet cave are not really horses. (Sapolsky)
- In recent years, scientists have made remarkable insights into the neurobiology of symbols. A major finding from their work is that the brain is not very good at distinguishing between the metaphorical and literal. In fact, symbols and metaphors, and the morality they engender, are the product of clunky processes in our brains. (Sapolsky)
- The best way to shine a light on this unwieldy process is through metaphors for two feelings critical to survival: pain and disgust. … There are fancier more recently evolved parts of the brain in the frontal cortex that assess the meaning of pain. Maybe [a pain signal is] bad news, or maybe it’s good news. Much of this assessing occurs in a frontal cortical region called the anterior cingulate. This structure is heavily involved in “error detection,” noting discrepancies between what is anticipated and what occurs. … In experimental settings, you’re playing with two [people] and suddenly they start ignoring you and only toss the ball between them. Junior high all over again. And the brain scanner shows that the neurons in your anterior cingulate activate. In other words, rejection hurts. … Both abstract social and literal pain impact the same cingulate neurons. (Sapolsky)
- We take things a step further. While in a brain scanner, you’re administered a mild shock, delivered through electrodes on your fingers. All the usual brain regions activate, including the anterior cingulate. Now you watch your beloved get shocked in the same way. The brain regions that ask, “Is it my finger or toe that hurts?” remain silent. It’s not their problem. But your anterior cingulate activates, and as far as it’s concerned, “feeling someone’s pain” isn’t just a figure of speech. You seem to feel the pain too. As evolution continued to tinker, it did something remarkable with humans. It duct-taped the anterior cingulate’s role in giving context to pain into a profound capacity for empathy. (Sapolsky)
- Studies show the human anterior cingulate is more complex than in other species, with more connections to abstract, associational parts of the cortex, regions that can call your attention to the pains of the world, rather than the pain in your big toe. (Sapolsky)
- Our brains’ shaky management of symbols adds tremendous power to a unique human quality: morality. You’re in a brain scanner and because of the scientist’s weirdly persuasive request, you bite into some rotten food. Something rancid and fetid and skanky. This activates another part of the frontal cortex, the insula, which, among other functions, processes gustatory and olfactory disgust. It sends neuronal signals to face muscles that reflexively spit out that bite, and to your stomach muscles that make you puke. All mammals have an insula that processes gustatory disgust. But we are the only animal where that process serves something more abstract. … Think about something awful you once did, something deeply shameful. The insula activates. It has been co-opted into processing that human invention: moral disgust. (Sapolsky)
Brief comments to close
Once again, let’s summarize all of the above research into a hierarchical chart. This one mimics the format of the one I produced for the functions of consciousness, but now we have its companion for the mechanisms as well.

Please note once again that these are the mechanisms of consciousness as they now exist in biotic life. As I have said before, there is nothing stopping artificial life from experiencing its own unique feelings of consciousness via new mechanisms. But I maintain that these are still the hierarchies that must be observed if artificial consciousness is become one that we will recognise.
Next up, I’ll tackle the ontogeny of consciousness in a human being. Hopefully that will fit just as well into this hierarchy and my use of Tinbergen’s 4 questions will continue to provide a comprehensive understanding of this complex phenomenon. I’ll bet you can’t wait!
--------------------------------------------
Previous Posts in This Series:
Consciousness 1 — Introduction to the Series
Consciousness 2 — The Illusory Self and a Fundamental Mystery
Consciousness 3 — The Hard Problem
Consciousness 4 — Panpsychist Problems With Consciousness
Consciousness 5 — Is It Just An Illusion?
Consciousness 6 — Introducing an Evolutionary Perspective
Consciousness 7 — More On Evolution
Consciousness 8 — Neurophilosophy
Consciousness 9 — Global Neuronal Workspace Theory
Consciousness 10 — Mind + Self
Consciousness 11 — Neurobiological Naturalism
Consciousness 12 — The Deep History of Ourselves
Consciousness 13 — (Rethinking) The Attention Schema
Consciousness 14 — Integrated Information Theory
Consciousness 15 — What is a Theory?
Consciousness 16 — A (sorta) Brief History of Its Definitions
Consciousness 17 — From Physics to Chemistry to Biology
Consciousness 18 — Tinbergen's Four Questions
Consciousness 19 — The Functions of Consciousness
Next up, I’ll tackle the ontogeny of consciousness in a human being. Hopefully that will fit just as well into this hierarchy and my use of Tinbergen’s 4 questions will continue to provide a comprehensive understanding of this complex phenomenon. I’ll bet you can’t wait!
--------------------------------------------
Previous Posts in This Series:
Consciousness 1 — Introduction to the Series
Consciousness 2 — The Illusory Self and a Fundamental Mystery
Consciousness 3 — The Hard Problem
Consciousness 4 — Panpsychist Problems With Consciousness
Consciousness 5 — Is It Just An Illusion?
Consciousness 6 — Introducing an Evolutionary Perspective
Consciousness 7 — More On Evolution
Consciousness 8 — Neurophilosophy
Consciousness 9 — Global Neuronal Workspace Theory
Consciousness 10 — Mind + Self
Consciousness 11 — Neurobiological Naturalism
Consciousness 12 — The Deep History of Ourselves
Consciousness 13 — (Rethinking) The Attention Schema
Consciousness 14 — Integrated Information Theory
Consciousness 15 — What is a Theory?
Consciousness 16 — A (sorta) Brief History of Its Definitions
Consciousness 17 — From Physics to Chemistry to Biology
Consciousness 18 — Tinbergen's Four Questions
Consciousness 19 — The Functions of Consciousness

